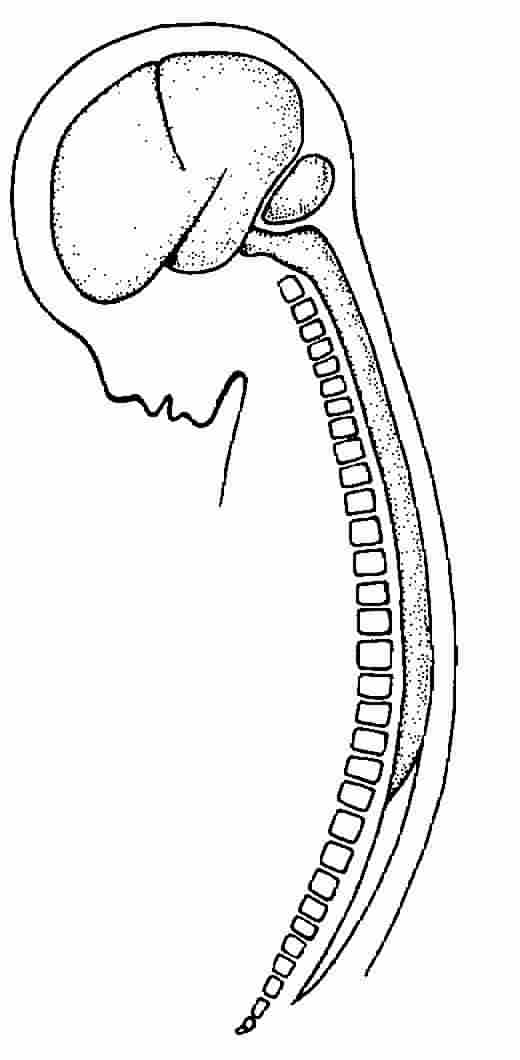
Men ought to know that from the brain, and from the brain
only, arise our pleasures, joys, laughter and jests, as well as our
sorrows, pains, griefs and tears. --Hippocrates (5th
Century, B.C.; quoted in Kandel et al. 1991:iv)
Neuro term. 1. Those circuits, centers, and
modules of the central nervous system involved in sending, receiving,
and processing speechless signs. 2. In right-handed individuals,
modules of the right brain cerebral hemisphere, considered to be more
nonverbal, holistic, visuospatial, and
intuitive than the verbal, analytic, sequential, and rational left
brain hemisphere (see HUMAN BRAIN, Right brain, left brain).
3. Those ancient centers (e.g., nuclei) and paleocircuits of the nervous system which evolved in
vertebrates--from the jawless fishes to human ancestors (e.g., Homo
habilis)--for communication before the advent of speech.
Usage: Just as the brain's newer speech centers (e.g., Broca's area) control language communication, earlier areas of the nonverbal brain control communication apart from words. Knowing its parts and wiring helps us decode nonverbal messages.
Central pattern generators (CPGs). Chief among the nervous system's parts and wiring are CPGs. CPGs are mini-networks of interneurons located in areas of the spinal cord and hindbrain that control rhythmic muscular movements such as breathing, chewing, hopping, jumping, running, scratching, shivering, swallowing, swimming, and walking. While such repeated actions may be initiated by cortical or emotional centers above, their rhythms are encoded, programmed, and controlled by the CPGs below. Found in chordates and invertebrates alike, CPGs are of recent discovery and incompletely understood (there is no mention of "central pattern generator," e.g., in my 38th edition desk copy of Gray's Anatomy [1995]).
Salience network. "The term 'salience network' refers to a suite of brain regions whose cortical hubs are the anterior cingulate and ventral anterior insular (i.e., frontoinsular) cortices. This network, which also includes nodes in the amygdala, hypothalamus, ventral striatum, thalamus, and specific brainstem nuclei, coactivates in response to diverse experimental tasks and conditions, suggesting a domain-general function. In the 12 years since its initial description, the salience network has been extensively studied, using diverse methods, concepts, and mammalian species, including healthy and diseased humans across the lifespan. Despite this large and growing body of research, the essential functions of the salience network remain uncertain"(William W. Seeley, Journal of Neuroscience, 11 Dec. 2019, pp. 9878-9882).
Media. "A skillful outline can be more appealing than a photographic
image. The simple line appeals to the brain which has limited attention and
limited abilities to process information rapidly." (San Diego
Union-Tribune interview with UC-San Diego neuroscientist, Vilayanur
Ramachandran [May 7, 1999, A1, A-19])
Literature. The first
recorded verbal reference to the human brain is Egyptian (the word
'ys), written on papyrus in the 17th Century, B.C. (Kandel et al.
1991).
Evolution. Our nonverbal brain consists of six
interrelated divisions, outlined below, which merged in an evolutionary process
from circa 500-to-two million years ago:
(1) AQUATIC BRAIN & SPINAL CORD: The oldest neural division, present in the jawless fishes, includes the spinal cord's interneuron pools and motor neuron pathways a. for tactile withdrawal, and b. for the rhythmic, oscillatory movements of swimming (and much later, for walking).
(2) AMPHIBIAN BRAIN: With amphibians, 1. the pontine reticular excitatory system becomes more elaborate. The pontine tegmentum's link to the spinal cord's anterior horn motor neurons and muscle spindles raised the body by exciting antigravity extensor muscles (see ANTIGRAVITY SIGN). 2. The vestibulospinal pathway elaborated--from receptors in the inner ear via the vestibular nerve (cranial VIII), and via cerebellar fibers to the vestibular nucleus in the upper medulla--running the length of the spinal cord for body posture (i.e., basic stance) in relation to gravity. 3. The tectospinal tract evolved, consisting of the superior (and inferior) colliculus and its links, via the brain stem, running a. to cervical cord interneurons, then b. to anterior horn motor neurons, then c. to spinal nerves, and finally reaching d. muscle spindles for postural reflexes to sights and sounds. 4. And the rubrospinal tract further evolved: paleocircuits from the red nucleus of the midbrain running a. to thoracic cord interneurons, then b. to anterior horn motor neurons, and finally c. to muscles and muscle spindles for postural tone of the limbs' flexor muscles.
(3) REPTILIAN BRAIN: With reptiles, 1. the vestibuloreticulospinal system evolved to control axial and girdle muscles for posture relative to positions of the head. 2. The basal ganglia-ansa lenticularis pathway reverberated links between the amygdala and basal ganglia via the ansa lenticularis and lenticulate fasciculus to the midbrain tegmentum, red nucleus, and reticular system to spinal cord interneurons required for the high-stand display.
(4) MAMMALIAN BRAIN: With mammals, 1. the amygdalo-hypothalamic tract became more elaborate: the central amygdala's link to the hypothalamus, via the stria terminalis, provided wiring for defensive postures (see, e.g., BROADSIDE DISPLAY). 2. Hypothalamus-spinal cord pathways adapted as well: the hypothalamus's dorsomedial and ventromedial nuclei fed a. indirectly via the brain stem's reticular system, and b. directly through fiberlinks to lower brain-stem and spinal-cord circuits to cord motor neurons for emotion cues (see, e.g., ANGER). 3. The septo-hypothalamo-midbrain continuum evolved: the medial forebrain bundle (from the olfactory forebrain and limbic system's septal nuclei) via the hypothalamus's lateral nuclei to midbrain-tegmentum brain-stem motor centers, mediated emotions (see, e.g., FEAR). 4. The cingulate gyrus facial circuit evolved: links run from the anterior cingulate cortex a. to the hippocampus, b. to the amygdala, c. to the hypothalamus, and d. through the brain stem, finally e. to the vagus (cranial X) and facial (cranial VII) nerves which, respectively, control the larynx and facial muscles required for vocalizing and moving the lips.
(5) PRIMATE BRAIN: With primates, 1. the neocortex's corticospinal tract further evolved: the posterior parietal cortex linked to supplementary motor, premotor, and primary motor cortices (with basal-ganglia feedback loops) via the corticospinal tract, to cervical and thoracic anterior-horn spinal interneurons, and to motor neurons in control of arm, hand, and finger muscles for skilled movements of the precision grip. 2. Modules of the inferior temporal neocortex evolved to provide visual input a. to the occipital neocortex's parvocellular interblob system (V1 to V2 and V4), permitting recognition of complex shapes, and b. to the inferior temporal cortex permitting heightened responses to hands and the ability to recognize faces.
(6) HUMAN BRAIN: With hominids, 1. the
corticobulbar tract further evolved: corticobulbar pathways to the
facial nerve (cranial VII) permitted intentional facial
expressions (see, e.g., SMILE). 2. Broca's cranial
pathways evolved: Broca's-area neocircuits via corticobulbar pathways to
multiple cranial nerves permitted speech.
3. And Broca's spinal pathways evolved: Broca's-area
neocircuits via corticospinal pathways to cervical and thoracic spinal nerves
permitted manual sign language and linguistic-like mime
cues.
Neuro-notes I. Mirror neurons: Mirror neurons enable us to read others' nonverbal body movements as if they were our own. Consider Vittorio Gallese's abstract for the 2012 conference on "Mirror Neurons: New Frontiers 20 Years After Their Discovery": " 'Embodied Simulation' (ES) provides a new empirically based notion of intersubjectivity, viewed first and foremost as intercorporeity. ES challenges the notion that Folk Psychology is the sole account of interpersonal understanding. Before and below mind reading is intercorporeity as the main source of knowledge we directly gather about others. By means of ES we do not just see an action, an emotion, or a sensation and then understand it through an inference by analogy. By means of ES we can map others' actions by re-using our own motor representations, as well as others, emotions and sensations by re-using our own viscero-motor and somatosensory representations. ES provides an original and unitary account of basic aspects of intersubjectivity, demonstrating how deeply our making sense of others living and acting bodies is rooted in the power of re-using our own motor, emotional and somatosensory resources." (See ISOPRAXISM, Neuro-notes IV; and New Frontiers in Mirror Neuron Research, edited by Pier Ferrari and Giacomo Rizzolatti, Oxford: Oxford U. Press, 2015.)
Neuro-notes II. Mirror neurons: ". . . we used to think that sensory systems and the motor systems [of the brain] are completely separate. This processing of sensory information [is such that] your movement [e.g., picking up a pencil] is [also] occurring in [the observer's] motor system. So a fraction of the cells [in the observer's brain] that are involved in picking up [a] pencil will respond when you pick up your pencil. It's in the motor system that he [Giacomo Rizzolatti] discovered this remarkable thing. So he made us realize that the motor systems have sensory cognitive capabilities. It's an extraordinary advance." (Source: Comments by Eric Kandel [with David Givens's edits, in brackets, for clarity] on PBS's "Charlie Rose Show" ("The Social Brain," January 10, 2010); http://www.charlierose.com/download/transcript/10820 [accessed December 19, 2012]; copyright 2010 Charlie Rose; transcription copyright 2010 CQ Transcriptions, LLC)
See also ENTERIC
BRAIN, NONVERBAL LEARNING, ORIENTING
REFLEX.
Copyright 1998 - 2022 (David B. Givens/Center for Nonverbal Studies)
Illustration credit: Principles of Neural Science, copyright 1991 by Appleton & Lange, Norwalk, Connecticut (Kandel et al. 1991)